The world of plants, with its silent, green complexity, often holds secrets that reshape our understanding of life itself. We know plants need light to grow, a fundamental truth ingrained from childhood. But what if light wasn’t just a simple switch, either on or off, but a symphony of wavelengths, each playing a crucial role, and some even enhancing the performance of others? This intricate dance of photons and pigments is precisely what the Emerson Effect illuminates. Discovered over half a century ago, this groundbreaking phenomenon revealed a synergistic interaction between different light wavelengths, forever changing how we perceive and utilize light for plant growth. It’s a tale of scientific discovery, photosynthetic marvels, and practical applications that continue to drive innovation in horticulture and agriculture.
The Discovery of Synergy: What is the Emerson Effect?
Before the mid-20th century, the prevailing belief was that the efficiency of photosynthesis was directly proportional to the amount of light absorbed by chlorophyll, peaking around specific wavelengths (red and blue light). However, the pioneering work of American plant physiologist Robert Emerson and his colleagues in the 1950s began to challenge this simplistic view, uncovering a remarkable complexity in how plants process light energy.
Emerson’s initial experiments involved measuring the quantum yield of photosynthesis – the number of oxygen molecules produced per absorbed photon – in the green alga Chlorella. He observed a curious phenomenon known as the “red drop.” When algae were illuminated with monochromatic light in the far-red region of the spectrum (wavelengths greater than approximately 680 nanometers), the quantum yield of photosynthesis significantly decreased. This was unexpected because chlorophyll still absorbed light in this region, albeit less efficiently than at peak red light absorption.
The pivotal discovery came when Emerson and his team provided a second, shorter wavelength of light (typically red light, around 650-680 nm) simultaneously with the far-red light. What they found was astonishing: the rate of photosynthesis under the combined illumination was significantly greater than the sum of the rates obtained when each wavelength was applied individually. This phenomenon, where two different wavelengths of light work together to produce a photosynthetic rate higher than the sum of their independent effects, is known as the Emerson Enhancement Effect or simply the Emerson Effect.
This “enhancement effect” was a clear indication of a synergistic interaction. It implied that the photosynthetic machinery was not a single, monolithic system, but rather comprised of at least two distinct photochemical systems that could be independently excited by different wavelengths and, more importantly, could cooperate to boost overall efficiency. This discovery laid the foundation for our modern understanding of how light energy is captured and converted into chemical energy within plant cells, fundamentally changing the scientific landscape of photosynthesis.
The Photosynthetic Machinery: Why Does Synergy Happen?
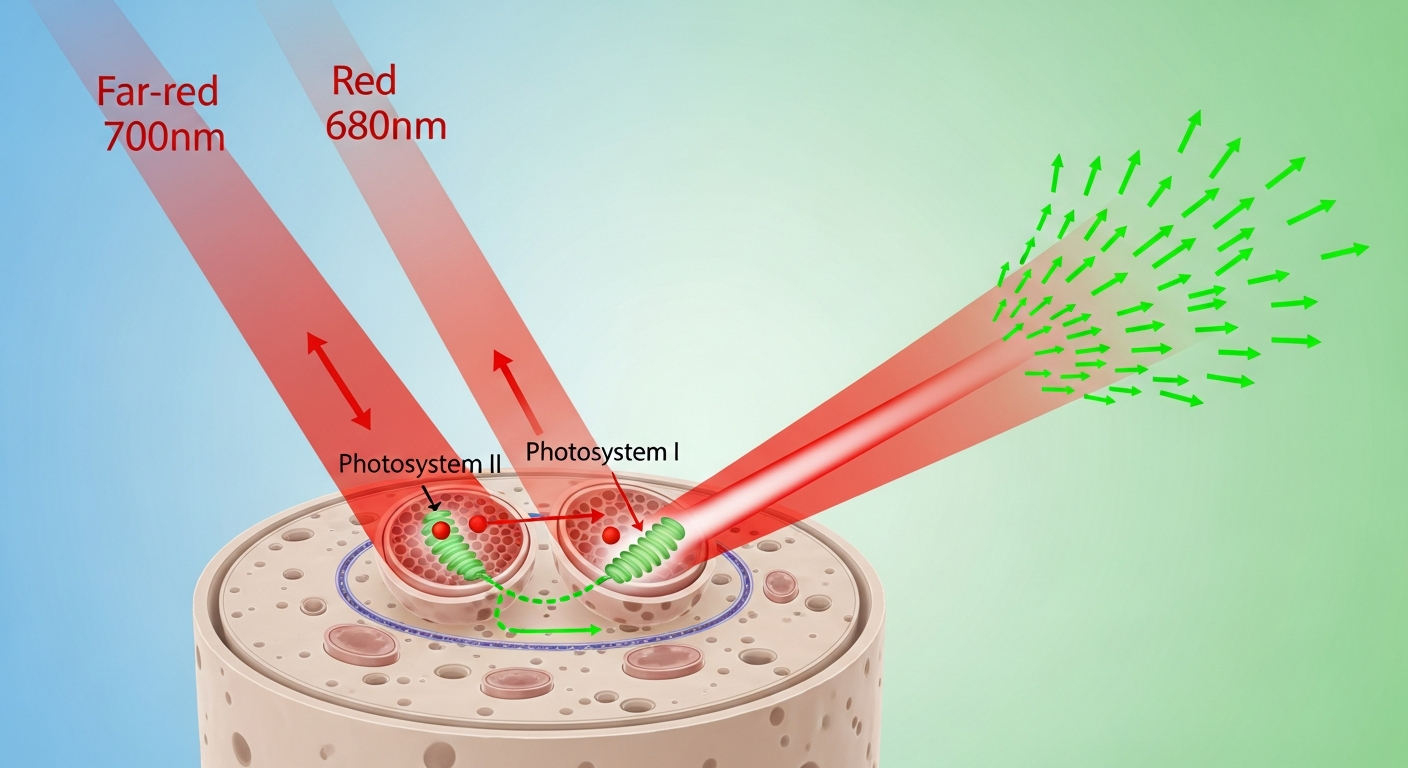
The existence of the Emerson Effect provided compelling evidence for the presence of multiple photosystems within chloroplasts, a concept that was further elucidated by subsequent research. Today, we understand that photosynthesis in plants and algae relies on two interconnected protein-pigment complexes: Photosystem I (PSI) and Photosystem II (PSII). These photosystems work in tandem, connected by an electron transport chain, to convert light energy into chemical energy in a process often referred to as the “Z-scheme.”
Photosystem II (PSII) is primarily responsible for the initial capture of light energy, specifically excelling at absorbing shorter wavelengths of light, particularly in the red spectrum (around 680 nanometers). When PSII absorbs light, it initiates the splitting of water molecules (photolysis), releasing electrons, protons, and oxygen as a byproduct. These electrons are then passed along an electron transport chain, contributing to the creation of a proton gradient that drives ATP synthesis.
Photosystem I (PSI), on the other hand, is optimized for absorbing slightly longer wavelengths, including far-red light (up to 700-730 nanometers), although it can also utilize shorter wavelengths. PSI receives electrons from the electron transport chain (which originated from PSII) and re-excites them with absorbed light energy. These energized electrons are then used to reduce NADP+ to NADPH, another crucial energy carrier for the synthesis of sugars.
The key to the Emerson Effect lies in the differential excitation and balancing of these two photosystems. When plants are exposed solely to far-red light (beyond ~680 nm), PSII activity significantly decreases because its primary pigments, like chlorophyll a-680, absorb very poorly at these wavelengths. This creates an imbalance: PSI might still be active, but it receives an insufficient supply of electrons from a “starved” PSII. The entire electron transport chain slows down, leading to the observed “red drop” in photosynthetic efficiency.
Conversely, if plants are exposed predominantly to red light around 680 nm, PSII can become highly excited, leading to an overabundance of electrons entering the electron transport chain. However, if there isn’t enough far-red light to adequately excite PSI and pull those electrons through, the electron transport chain can become “backed up.” This can lead to inefficient energy transfer and even potential damage from excess excitation energy.
The synergistic interaction observed in the Emerson Effect occurs because the simultaneous application of both red and far-red light provides the optimal excitation for both photosystems. Red light efficiently activates PSII, ensuring a robust supply of electrons, while far-red light effectively activates PSI, efficiently pulling those electrons through the electron transport chain and preventing bottlenecks. This balanced excitation of both PSI and PSII ensures a smooth, rapid, and highly efficient flow of electrons, maximizing the overall quantum yield of photosynthesis. It’s like having two pumps working in series; if one isn’t receiving enough input or isn’t able to pump fast enough, the whole system slows down. Providing the right “fuel” (wavelengths) to both pumps at the same time optimizes the entire process.
Beyond the Basics: Implications and Nuances of the Emerson Effect
The initial discovery of the Emerson Effect was a monumental step, but subsequent research has deepened our understanding of its nuances and broader implications, revealing that the interplay of light goes far beyond simple energy capture.
The “Red Drop” and Action Spectrum
The “red drop” phenomenon, the sharp decrease in photosynthetic efficiency at wavelengths greater than 680 nm, was the initial puzzle that led to the Emerson Effect’s discovery. It highlighted that the overall action spectrum of photosynthesis—the rate of a physiological activity plotted against wavelength—is not simply identical to the absorption spectrum of chlorophyll. While chlorophylls absorb strongly in the red and blue regions, the efficiency drops off in the far-red unless complemented by shorter red wavelengths. This distinction underscores that the entire photosynthetic apparatus, not just individual pigments, determines the efficiency of light utilization.
Different Plant Species and Environmental Factors
The magnitude and optimal wavelength ratios for the Emerson Effect can vary significantly across different plant species. For instance, some deep-water aquatic plants or plants growing in shaded environments might have evolved mechanisms to utilize far-red light more effectively, as it penetrates deeper into water or dense canopies. The specific balance between PSI and PSII activity, and thus the extent of the enhancement effect, can also be influenced by various environmental factors such as light intensity, temperature, CO2 concentration, and even nutrient availability. For example, under very high light intensities, the electron transport chain might already be saturated, potentially reducing the relative enhancement from far-red light, or conversely, making the balance even more critical to prevent photoinhibition.
The Role of Far-Red Light Beyond Enhancement
While historically seen primarily as an enhancer for red light in photosynthesis, recent research has revealed that far-red light plays several independent and crucial roles in plant development, collectively known as photomorphogenesis. Plants use specialized photoreceptors, primarily phytochromes, to sense the ratio of red to far-red light. This ratio acts as a powerful environmental signal:
- Shade Avoidance Syndrome: In dense canopies, plants perceive a lower red:far-red ratio because chlorophyll in overlying leaves absorbs red light more strongly than far-red light. This signals competition, triggering a “shade avoidance syndrome” characterized by stem elongation, upward leaf orientation, and reduced branching, allowing the plant to reach for light.
- Flowering and Dormancy: The red:far-red ratio influences the timing of flowering in many species, a critical aspect of plant reproduction. It also plays a role in the induction and breaking of dormancy.
- Leaf Expansion and Chloroplast Development: Far-red light can influence leaf size and the development of chloroplasts, affecting overall photosynthetic capacity.
- Overall Biomass and Yield: Studies show that adding far-red light to an otherwise complete spectrum can independently increase biomass and yield in many crops, even beyond its synergistic effect on instantaneous photosynthetic rates. This suggests that its developmental roles contribute significantly to final plant size and productivity.
Therefore, far-red light is not merely a supplementary wavelength; it’s a vital component of the plant’s light environment, influencing both its immediate energy conversion processes and its long-term growth and development.
Harnessing the Emerson Effect: Practical Applications in Horticulture
The scientific understanding of the Emerson Effect, coupled with advancements in lighting technology, has profound implications for modern horticulture and controlled environment agriculture. Growers are increasingly moving beyond simply providing “enough” light to providing the “right” kind of light, precisely tailored to plant needs.
Optimizing Grow Light Spectrums
The advent of LED grow lights has revolutionized our ability to precisely control the spectral output of artificial illumination. Unlike older technologies like HPS or MH lamps that offered broad, fixed spectrums, LEDs can be configured to emit specific wavelengths, allowing growers to fine-tune the light environment. This capability makes harnessing the Emerson Effect highly practical:
- Balanced Red and Far-Red Ratios: Modern LED fixtures often incorporate specific red (e.g., 660 nm) and far-red (e.g., 730 nm) diodes, alongside blue and other wavelengths. Growers can adjust the intensity and ratio of these wavelengths to maximize photosynthetic efficiency and trigger desired photomorphogenetic responses.
- Moving to Tuned Spectrums: Instead of relying on a generic “full spectrum,” advanced growers are developing “tuned spectrums” that are specifically designed for different plant species, growth stages (e.g., vegetative, flowering), and even desired crop qualities. Incorporating far-red light is now recognized as a critical component of these optimized spectrums.
- Consideration of Plant Stage: While vegetative growth often benefits from a good balance of red and blue, the flowering stage in many plants can be significantly enhanced by the addition of far-red light, impacting flower size, density, and secondary metabolite production.
Enhancing Crop Yield and Quality
By applying the principles of the Emerson Effect, growers can achieve significant improvements in crop production:
- Increased Biomass and Growth Rates: The enhanced photosynthetic efficiency directly translates to faster growth and greater overall biomass accumulation. This means quicker crop cycles and higher yields per square foot.
- Improved Product Quality: In many horticultural crops, particularly leafy greens, herbs, and cannabis, the inclusion of far-red light can positively influence attributes like leaf expansion, chlorophyll content, nutrient uptake, and the production of beneficial secondary metabolites (e.g., terpenes, cannabinoids, antioxidants). This leads to higher-quality, more marketable produce.
- Energy Efficiency: By making photosynthesis more efficient, growers can achieve higher yields with the same or even reduced electrical input for lighting, improving the energy footprint and profitability of indoor farms and greenhouses.
Tips for Growers to Leverage Far-Red Light and the Emerson Effect
For those looking to optimize their growing operations, here’s practical advice:
- Invest in Quality LED Fixtures: Choose grow lights that offer a broad spectrum, including dedicated far-red diodes, or fixtures with adjustable spectral output. Avoid “blurple” lights that lack comprehensive spectral control.
- Understand Your Crop’s Specific Needs: Research the ideal light spectrum and red:far-red ratios for the specific plants you are growing. Different species have different optimal responses.
- Monitor and Experiment: Start with recommended spectrums and then carefully observe plant response. Look for changes in growth rate, internode spacing, leaf size, and overall vigor. Small adjustments to the red:far-red ratio can yield noticeable results.
- Consider Far-Red as an “Add-on”: Some growers use supplemental far-red bars to complement existing full-spectrum lights, particularly during specific growth phases or to induce desired morphological changes like flowering.
- Integrate with Other Environmental Controls: Light spectrum is just one piece of the puzzle. Ensure optimal temperature, humidity, CO2 levels, and nutrient delivery to fully capitalize on enhanced photosynthetic efficiency.
- Embrace the “End-of-Day” Treatment: In some cases, a short burst of far-red light at the end of the day can help plants transition into the dark period more efficiently, promoting earlier flowering and influencing sleep cycles (nyctinasty).
Conclusion: The Unfolding Story of Light and Life
The Emerson Effect stands as a testament to the profound complexity and elegant efficiency of nature’s life-sustaining processes. What began as a puzzling observation by Robert Emerson over 60 years ago blossomed into a foundational principle of plant physiology. It revealed that photosynthesis is not merely about absorbing light, but about orchestrating a delicate balance between different light-capturing systems, a synergistic interaction that maximizes energy conversion and underpins nearly all life on Earth.
Today, our understanding of the Emerson Effect and the multifaceted roles of different light wavelengths continues to evolve. From unraveling the intricate molecular mechanisms within chloroplasts to pioneering advanced horticultural lighting solutions, this discovery remains a cornerstone of plant science. As we push the boundaries of controlled environment agriculture, indoor farming, and sustainable food production, the ability to precisely manipulate the light spectrum, guided by the principles of synergistic wavelength interaction, will be increasingly critical. The story of light and life is far from over; the Emerson Effect is just one brilliant chapter in a narrative that continues to inspire innovation and deepen our awe for the natural world.