For decades, the world of horticulture and plant science operated under a seemingly simple yet foundational concept: Photosynthetically Active Radiation, or PAR. Defined as the spectral range from 400 to 700 nanometers (nm), PAR represented the wavelengths of light that plants primarily use for photosynthesis. It was the golden standard, guiding greenhouse design, grow light technology, and agricultural practices. However, as our understanding of plant biology deepens and artificial lighting technologies, particularly LEDs, become ever more sophisticated, a paradigm shift is underway. Scientists and growers are now exploring the critical role of light beyond the traditional PAR range, venturing into the realm of far-red light and embracing the concept of extended Photosynthetically Active Radiation, or ePAR. This evolution is not merely a technical adjustment; it’s a profound re-evaluation of how plants perceive and utilize light, promising unprecedented control over plant growth, development, and yield in modern agriculture.
Beyond PAR: Unpacking the Traditional Photosynthetically Active Radiation (PAR) Concept
To truly appreciate the significance of ePAR and far-red light, it’s essential to first understand the traditional definition and limitations of Photosynthetically Active Radiation (PAR). Historically, PAR was defined as the portion of the electromagnetic spectrum spanning from 400 nm (blue light) to 700 nm (red light). This definition was largely based on early measurements of photosynthetic efficiency, which showed that light within this range was most effectively absorbed by chlorophyll pigments, the primary light-harvesting molecules in plants.
Chlorophyll a and chlorophyll b, the two main types of chlorophyll, exhibit strong absorption peaks in the blue (around 430-470 nm) and red (around 640-670 nm) regions of the spectrum. These absorption characteristics led to the long-held belief that these wavelengths were the most crucial for driving photosynthesis. Green light (around 500-600 nm) was often considered less important, as it is largely reflected by chlorophyll, giving plants their characteristic green appearance. Similarly, wavelengths outside the 400-700 nm range, including ultraviolet (UV) and far-red light, were traditionally deemed either detrimental or photo-biologically inactive for photosynthesis.
For many years, this PAR definition served as a practical benchmark for measuring light intensity for plant growth. Scientists and growers used quantum sensors that measured the Photosynthetic Photon Flux Density (PPFD), representing the number of photons per second per unit area within the 400-700 nm range. This metric proved adequate when dealing with sunlight, which provides a broad spectrum of light, or traditional grow lights like high-pressure sodium (HPS) and metal halide (MH), which also emit relatively broad and somewhat inefficient spectra. However, as horticulture moved into controlled environment agriculture (CEA) and embraced highly efficient, spectrally tunable LED grow lights, the limitations of the traditional PAR concept became increasingly apparent. The ability to precisely dial in specific wavelengths of light revealed that plants are far more sophisticated in their light perception and utilization than previously assumed, particularly concerning the often-overlooked far-red spectrum.
The Far-Red Frontier: Introducing ePAR and its Scientific Basis
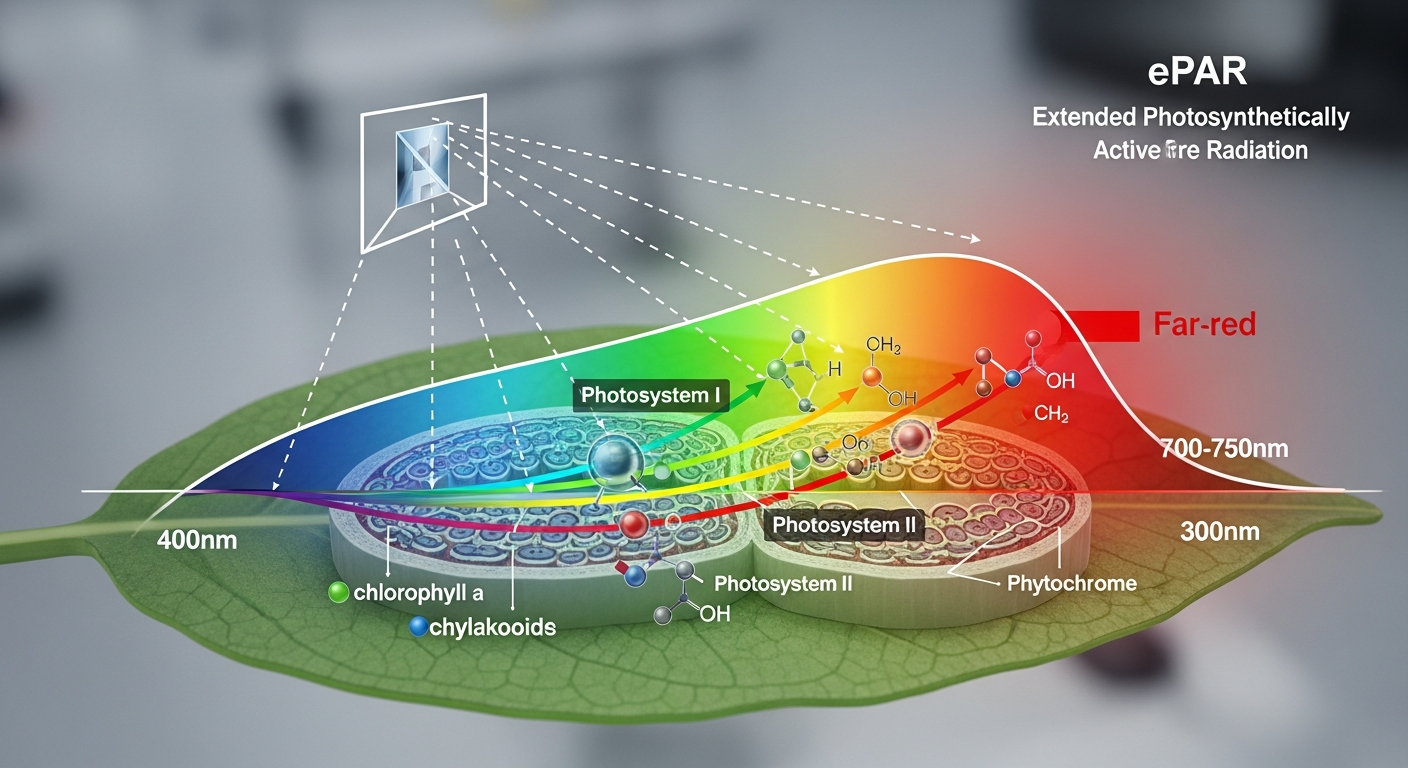
The emergence of advanced research and lighting technology has led to a significant expansion of our understanding of photosynthetically active radiation, culminating in the concept of ePAR, or extended Photosynthetically Active Radiation. While traditional PAR encompasses 400-700 nm, ePAR typically extends this range to include far-red light, often up to 750 nm, and in some studies, even up to 800 nm. This shift acknowledges that far-red light, once thought to be minimally photosynthetically active, plays a crucial and multifaceted role in plant growth and development.
The scientific basis for ePAR and the inclusion of far-red light is rooted in several key discoveries and principles of plant physiology:
The Emerson Effect and Quantum Yield
One of the most foundational pieces of evidence for the photosynthetic activity of far-red light comes from the “Emerson Enhancement Effect,” discovered by Robert Emerson in the 1950s. Emerson observed that photosynthetic efficiency significantly increased when algae were exposed to a combination of red light (around 680 nm) and far-red light (around 700 nm), compared to the sum of efficiencies when exposed to each wavelength individually. This synergistic action, known as the Emerson effect, demonstrated that two distinct photosystems (Photosystem I and Photosystem II) operate in concert during photosynthesis.
Photosystem II (PSII) is primarily responsible for absorbing shorter wavelengths of light (blue and red up to about 680 nm) and splitting water molecules. Photosystem I (PSI), on the other hand, can effectively absorb longer wavelengths, including far-red light (up to around 750 nm). While PSI can perform cyclic electron flow on its own, it is much more efficient when paired with PSII. When red light (activating PSII) and far-red light (activating PSI) are provided simultaneously, the electron transport chain is balanced and optimized, leading to a higher overall quantum yield of photosynthesis – meaning more sugar produced per photon absorbed. This improved balance in the electron transport chain contributes to increased biomass accumulation and greater energy efficiency, especially under high light intensities where electron flow can become imbalanced.
The Photoreceptor Symphony: Phytochrome and Cryptochromes
Beyond direct photosynthetic energy conversion, far-red light profoundly influences plant development through its interaction with specialized photoreceptors. The most significant of these is phytochrome, a remarkable protein pigment that acts as a primary light sensor in plants. Phytochrome exists in two interconvertible forms: Pr (phytochrome red-absorbing) and Pfr (phytochrome far-red-absorbing).
-
Pr form: This is the inactive form of phytochrome, which primarily absorbs red light (around 660 nm). Upon absorbing red light, Pr rapidly converts to the Pfr form.
-
Pfr form: This is the biologically active form of phytochrome. It absorbs far-red light (around 730 nm) and, upon doing so, converts back to the inactive Pr form. In the absence of far-red light, Pfr slowly reverts to Pr in darkness.
The ratio of red light to far-red light (R:FR ratio) in a plant’s environment dictates the proportion of Pr to Pfr. This ratio acts as a critical signal, influencing a vast array of developmental processes known as photomorphogenesis, including seed germination, seedling de-etiolation, stem elongation, leaf expansion, flowering time, and shade avoidance response. A high R:FR ratio (more red light) indicates ample light availability and promotes compact growth, while a low R:FR ratio (more far-red light, as found under a canopy) signals competition for light, triggering shade avoidance responses.
While phytochrome is the primary sensor for red and far-red light, it’s also important to acknowledge other photoreceptors like cryptochromes and phototropins, which primarily sense blue light and regulate processes like hypocotyl elongation, stomatal opening, and phototropism. However, for understanding the unique influence of far-red light, phytochrome remains the star player, orchestrating many of the intricate developmental changes observed in plants.
Far-Red’s Profound Impact on Plant Physiology and Development
The integration of far-red light into our understanding of photosynthesis and plant development has unveiled a spectrum of physiological responses that can be strategically harnessed in modern horticulture. Far-red light is far from inert; it is a powerful regulator, driving changes in morphology, biomass accumulation, and reproductive timing.
Enhanced Biomass Accumulation and Leaf Expansion
One of the most well-documented effects of including far-red light in the growing spectrum is a significant increase in overall plant biomass. This is often attributed to the combination of two factors:
-
Increased Photosynthetic Efficiency (Emerson Effect): As discussed, the synergistic action of far-red with red light optimizes the electron transport chain, leading to a higher quantum yield of photosynthesis. This means plants can convert light energy into chemical energy more efficiently, fueling greater growth.
-
Leaf Expansion: Far-red light actively promotes the expansion of individual leaves and overall leaf area. Larger leaves mean more surface area to capture incoming light, translating to greater photosynthetic capacity for the entire plant. While this can be beneficial for maximizing yield, it can also lead to thinner leaves and increased internode length, which might be undesirable for certain crops or growing systems.
In crops like lettuce and other leafy greens, a moderate amount of far-red can significantly boost marketable yield by increasing leaf size and weight. However, growers must balance this with desired plant architecture, as excessive far-red can lead to undesirable stretching.
Accelerated Flowering and Fruiting
For many flowering plants, far-red light plays a pivotal role in regulating the timing of floral initiation and subsequent fruit development. Phytochrome, through its R:FR sensing mechanism, is a key component of the photoperiodic clock, which dictates when a plant will transition from vegetative growth to reproductive growth.
-
Flowering Induction: In many long-day plants, exposure to far-red light can accelerate flowering, effectively shortening the time to harvest. Conversely, in short-day plants, far-red during the “dark period” (or what the plant perceives as darkness) can inhibit flowering, by effectively shortening the perceived dark period. Understanding these species-specific responses is crucial for spectral tuning.
-
Fruit Development and Quality: Beyond merely triggering flowering, far-red can also influence the development and quality of fruits. Research suggests that far-red can impact fruit size, sugar content, and even pigment development in some species, though these effects are highly crop-dependent and an active area of research.
Shade Avoidance Response (SAR)
One of the most striking and ecologically significant roles of far-red light is its involvement in the shade avoidance response (SAR). Plants in dense stands or under a canopy of other plants experience a shift in the quality of light reaching them. Chlorophyll in the upper leaves absorbs much of the red and blue light, while far-red light is transmitted or reflected. This results in a lower R:FR ratio reaching the lower leaves or shaded plants.
Plants detect this low R:FR ratio via their phytochrome system, interpreting it as a signal of impending competition for light. In response, they initiate a series of changes characteristic of SAR:
-
Stem Elongation: Plants “stretch” or elongate their stems and petioles to outcompete neighbors and reach higher light levels.
-
Reduced Branching: Energy is redirected from lateral branching to main stem growth.
-
Early Flowering: Some plants may flower earlier to complete their life cycle before being completely shaded out.
-
Changes in Leaf Angle: Leaves may orient themselves more vertically to minimize self-shading.
While beneficial in natural environments, SAR can be detrimental in controlled environment agriculture, leading to “leggy” plants, reduced yield quality, and wasted space. Growers must carefully manage the R:FR ratio to mitigate unwanted shade avoidance while still leveraging far-red’s benefits for biomass and photosynthetic efficiency.
Photosynthetic Efficiency and Stress Tolerance
Beyond the Emerson effect, far-red light contributes to the overall photosynthetic machinery by maintaining the balance of electron transport. By specifically activating Photosystem I, far-red can help prevent photoinhibition and mitigate stress under high light intensities, ensuring that the photosynthetic apparatus operates smoothly and efficiently. While research is ongoing, some studies also suggest that specific spectral compositions, including far-red, might play a role in enhancing plant resilience to certain environmental stressors, potentially through signaling pathways that regulate defense mechanisms.
Harnessing the Power of Far-Red: Practical Applications in Modern Horticulture
The scientific understanding of ePAR and the profound influence of far-red light has moved beyond academic curiosity to become a critical component of advanced horticultural practices. Modern growers in controlled environments now have the unprecedented ability to precisely tune light spectra, offering new levers to optimize plant growth, yield, and quality.
Optimizing LED Grow Light Spectrums for ePAR
The advent of Light Emitting Diodes (LEDs) has revolutionized horticultural lighting. Unlike traditional broad-spectrum sources, LEDs can be engineered to emit light at very specific wavelengths. This precision is what makes spectral tuning, and thus effective far-red integration, possible.
-
Moving Beyond Red/Blue: Early LED grow lights focused predominantly on red and blue wavelengths, aiming for maximum chlorophyll absorption. While effective, this approach often overlooked the regulatory and synergistic power of far-red.
-
Incorporating Far-Red Diodes: Modern, high-performance LED grow lights now routinely include dedicated far-red diodes (typically around 730 nm) as part of their spectral output. These are crucial for unlocking the Emerson effect and phytochrome-mediated responses.
-
Balancing the Spectrum: The key isn’t simply adding far-red, but integrating it effectively. The ideal ePAR spectrum is a balance of blue, green, red, and far-red light. The specific ratios will depend on the crop, growth stage, and desired physiological outcome.
-
Practical Tip: When selecting LED grow lights, look beyond just PPFD (400-700nm). Inquire about the Photosynthetic Photon Flux Density (ePPFD) which includes far-red (e.g., 400-750nm or 400-800nm) and review the spectral power distribution (SPD) graph to ensure adequate far-red presence. Consider fixtures with tunable far-red channels for maximum flexibility.
Strategic Far-Red Application for Specific Crop Goals
Understanding the effects of far-red allows growers to apply it strategically to achieve specific objectives:
-
For Increased Biomass (e.g., Leafy Greens): A moderate inclusion of far-red throughout the vegetative stage can promote larger leaves and overall plant size, leading to higher yields for crops like lettuce, basil, and herbs. However, monitor for excessive stretching.
-
For Accelerated Flowering (e.g., Cannabis, Ornamentals): For many long-day crops and certain short-day crops, a boost of far-red during specific photoperiods or at the end of the day (end-of-day far-red treatment) can significantly shorten the time to flowering, leading to faster crop cycles and increased throughput.
-
For Managing Plant Architecture: If excessive stretching (shade avoidance) is an issue, reducing the R:FR ratio or providing targeted far-red “pulses” can help mitigate this while still benefiting from increased biomass. Conversely, if some elongation is desired, for example, to increase internode spacing for better airflow, a higher far-red component can be beneficial.
-
For Propagation: Research suggests that far-red can sometimes promote rooting in cuttings, making it a valuable tool in propagation stages, though this is species-specific.
-
Practical Tip: Experimentation is key. Start with manufacturer recommendations for your crop type and then adjust far-red intensity or duration. For flowering crops, consider an “end-of-day far-red spike” – a brief period (15-30 minutes) of pure far-red light after the main lights turn off to rapidly convert residual Pfr to Pr, effectively extending the perceived dark period and accelerating flowering for certain species.
Measuring and Monitoring ePAR
As the industry embraces ePAR, traditional PAR meters that only measure 400-700 nm become insufficient. Accurate measurement and monitoring are crucial for effective spectral tuning.
-
ePPFD Meters: Growers need quantum sensors capable of measuring the extended PAR range, typically 400-750 nm or 400-800 nm, to get a true representation of the photosynthetically and morphogenetically active light environment.
-
Spectroradiometers: For advanced users, a spectroradiometer provides a complete spectral power distribution (SPD) graph, showing the intensity at every wavelength. This allows for precise calculation of ratios (e.g., R:FR ratio) and a deeper understanding of the light environment.
-
Practical Tip: If investing in new light measurement equipment, ensure it is calibrated to measure the extended PAR range. Understand the difference between PPFD (400-700nm) and ePPFD (e.g., 400-750nm) and apply the appropriate metric when evaluating light intensity for your cultivation strategy.
By understanding and strategically employing far-red light within an ePAR framework, modern growers can move beyond simply providing “enough light” to precisely tailoring the light environment to elicit specific, desired plant responses, leading to healthier plants, higher yields, and improved crop quality.
The journey from traditional PAR to the sophisticated understanding of ePAR and the critical role of far-red light represents a pivotal advancement in plant science and controlled environment agriculture. What was once dismissed as inefficient or irrelevant radiation is now recognized as a potent physiological signal and a direct contributor to photosynthetic efficiency. The Emerson effect elegantly demonstrates far-red’s ability to boost quantum yield, while the intricate dance between red and far-red light, mediated by the phytochrome photoreceptor system, orchestrates a symphony of developmental processes, from leaf expansion and biomass accumulation to flowering and the strategic avoidance of shade. As LED technology continues to evolve, offering unparalleled spectral control, the ability to precisely dial in optimal ePAR spectra, including specific far-red components, empowers growers to move beyond generic lighting solutions. This deeper understanding not only enhances productivity and resource efficiency but also opens new avenues for genetic expression and crop improvement, ushering in an exciting era where light is not just energy, but a powerful, finely tuned language we can now speak to our plants.